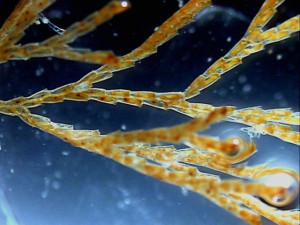
Description
Bugula neritina was described by Linnaeus from Europe in 1758. The "Bugula neritina" widely identified in fouling communities is apparently a widespread species complex of unknown tropical-warm-temperate origin, introduced to higher-latitude and more isolated ocean regions (Ryland 1960; Cohen and Carlton 1995). Biochemical studies indicate the presence of at least two cryptic species in California (Davidson and Haygood 1999). Field studies in Australia and North America show considerable variation in life history in "B. neritina" from different habitats, apparently due to genetic or early environmental effects (Keough 1989; Keough and Ross 1999).
Molecular studies of Bugula "neritina", and studies of its bacterial endosymbionts indicate that at least 3 genotypes occur in US waters, a type "D" found in deep waters off California, and presumed native to the northeastern Pacific (Davidson and Haygood 1999); a type "S" found in shallow waters of California (Davidson and Haygood 1999), also found from Cape Hatteras to the Gulf of Mexico and Caribbean, Hawaii, Australia, New Zealand, and England (McGovern and Hellberg 2003; Mackie et al. 2006), and a "North Atlantic" lineage, identified by specimens from DE, CT, and MA (McGovern and Hellberg 2003).
Taxonomy
| Kingdom | Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|---|
| Animalia | Bryozoa | Gymnolaemata | Cheilostomata | Bugulidae | Bugula neritina |
Synonyms
Invasion History
Chesapeake Bay Status
| First Record | Population | Range | Introduction | Residency | Source Region | Native Region | Vectors |
|---|---|---|---|---|---|---|---|
| 2000 | Established | Expanding | Introduced | Regular Resident | Western Atlantic | Unknown-Marine | Shipping(Fouling Community) |
History of Spread
Bugula neritina was described by Linnaeus from Europe in 1758, from the Mediterranean Sea. The name 'Bugula neritina', widely reported in fouling communities, applies to a widespread species complex, including species of unknown tropical-warm-temperate origin, now widespread and cryptogenic in equatorial regions, as well as locally distributed native forms. The cosmopolitan warm-water form (genotype S) has been introduced to higher-latitude and more isolated ocean regions (Ryland 1960; Cohen and Carlton 1995), including: the coast of northern Europe (1st record 1912- England, Netherlands, Belgium- Reise et al. 1999; Ryland 1960 ), the southwest Atlantic (1st record by 1937- Brazil-Argentina- Marcus 1937; Orensanz et al. 2002), the northeast Pacific (1st record 1904, southern CA-Puget Sound WA- Carlton 1979; Cohen and Carlton 1995), Hawaii (1st record 1921- Coles et al. 1999), and the southwest Pacific (Australia- by 1881- Keough and Ross 1999; New Zealand- by 1949- Cranfield et al. 1998). In the English Channel and Long Island Sound, it has appeared first near thermal effluents and gradually spread to other habitats (Carlton, personal communication 2004; Ryland 1960). In both the northeast Pacific and northwest Atlantic,B. "neritina" has been extending its range northward, reaching Puget Sound by 1994 (Cohen and Carlton 1995), and the Gulf of Maine (Boston and Salem Harbors, MA), by 2000 (MIT Sea Grant 2004).
On the Atlantic Coast of North America, Bugula "neritina" was identified by A. E. Verrill and reported by Coues and Yarrow (1878) from Fort Macon, near Beaufort NC. It was collected from the Dry Tortugas FL (Caribbean-Gulf of Mexico) by Osburn (1900), and appears to have been widespread in the Atlantic south of Cape Hatteras through the Gulf of Mexico, and through the Caribbean Sea by the early-mid 20th century (Maturo 1957; Osburn 1914, Osburn 1940; Osburn 1947; Shier 1964; Winston 1977). We regard this bryozoan as cryptogenic in the Atlantic south of Cape Hatteras and in the Caribbean and Gulf of Mexico.
Molecular studies of Bugula "neritina", and studies of its bacterial endosymbionts indicate that at least 2 genotypes occur in the northwest Atlantic ; a type "S" found from Cape Hatteras to the Gulf of Mexico and Caribbean, and also in Hawaii, Australia, New Zealand, and England (McGovern and Hellberg 2003; Mackie et al. 2006), and a "North Atlantic" lineage, identified by specimens from DE, CT, and MA (McGovern and Hellberg 2003). Type S appears to have been widely introduced around the world (Mackie et al. 2006), but its origin and the extent of its orignal range is unknown.
While the "North Atlantic lineage" of Bugula "neritina" appears to be genetically distinct from the widely introduced type "S", the history of B. "neritina", north of Cape Hatteras, strongly suggests a recent introduction. However, the orgin of this genotype is unknown. We predict that when more populations are studied, disjunct populations of the "North Atlantic lineage" will be found. Records are summarized here from north to south:
Chesapeake Bay region- In 2000 and 2001,"Bugula neritina" was collected on settling plates at Kiptopeke VA, on the Eastern shore at the mouth of the Bay (Ruiz et al., unpublished data). It was not collected at the mouth of the Bay, or in the adjacent Atlantic coastal bays by Osburn (1932, 1944).
Delaware Coastal Bays- McGovern and Hellberg (2003) obtained specimens for genetic studies from Indian River Bay DE.
Long Island Sound- 1985. "Bugula neritina" was first collected in 1985 in fouling at Millstone Nuclear Power Plant, in Waterford CT, in thermal effluents (Carlton, personal communication, 1999). In later years, this bryozoan became widespread at Avery Point, Groton CT, and other Long Island Sound locations away from thermal effluents (Osman, personal communication 2004).
Narragansett Bay- "Bugula neritina" was collected in 2000 at Coasters Harbor Island and Newport Shipyard at Newport RI, in the East Passage of Narragansett Bay. (MIT Sea Grant 2004).
Buzzards Bay-Vineyard Sound- McGovern and Hellberg (2003) obtained specimens for genetic studies from Woods Hole MA.
Gulf of Maine- "Bugula neritina" was collected in 2000 in Boston MA, at Black Falcon Terminal, Boston Harbor, at Salem MA in Salem Harbor and in 2003 at Brewer South Freeport Marina, Freeport ME in Casco Bay (MIT Sea Grant 2004).
History References- Carlton 1979; Carlton, personal communication 1999; Cohen and Carlton 1995; Coles et al. 1999; Coues and Yarrow 1878; Cranfield et al. 1998; Keough and Ross 1999; Marcus 1937; Maturo 1957; McGovern and Hellberg (2003); MIT Sea Grant 2004; Osburn 1900; Orensanz et al. 2002; Osburn 1914, Osburn 1932; Osburn 1940; Osburn 1944; Osburn 1947; Osman, personal communication 2004; Reise et al. 1999; Ruiz et al., unpublished data; Ryland 1960; Shier 1964; Winston 1977
Invasion Comments
Invasion Status, Vectors- Judy Winston (personal communication 2000) notes that there are a number of scattered records of B. neritina north of Cape Hatteras. She considers that these could represent shipping transport or Gulf Stream dispersal. Jim Carlton is skeptical of "natural" Gulf Stream transport of coastal, inshore fouling organisms, since none have been reported in extensive collecting of Gulf Stream Sargassum communities. (Carlton, personal communication 2004).
Ecology
Environmental Tolerances
| For Survival | For Reproduction | |||
|---|---|---|---|---|
| Minimum | Maximum | Minimum | Maximum | |
| Temperature (ºC) | 4.0 | 30.0 | ||
| Salinity (‰) | 18.0 | 40.0 | 20.0 | |
| Oxygen | None | |||
| pH | ||||
| Salinity Range | poly-eu |
Age and Growth
| Male | Female | |
|---|---|---|
| Minimum Adult Size (mm) | ||
| Typical Adult Size (mm) | 80.0 | 80.0 |
| Maximum Adult Size (mm) | 98.0 | 98.0 |
| Maximum Longevity (yrs) | ||
| Typical Longevity (yrs |
Reproduction
| Start | Peak | End | |
|---|---|---|---|
| Reproductive Season | |||
| Typical Number of Young Per Reproductive Event |
|||
| Sexuality Mode(s) | |||
| Mode(s) of Asexual Reproduction |
|||
| Fertilization Type(s) | |||
| More than One Reproduction Event per Year |
|||
| Reproductive Startegy | |||
| Egg/Seed Form |
Impacts
Economic Impacts in Chesapeake Bay
Bugula neritina is currently rare in Chesapeake Bay, and probably has no detectable economic impacts.
Economic Impacts Outside of Chesapeake Bay
Bugula neritina is a frequent fouling organism of fixed structures, ship hulls, in ships' internal water systems, and in powerplants using seawater in warm waters worldwide (Ryland 1971). Bugula neritina, collected in Botany Bay, Australia, proved highly resistant as larvae and adults to dissolved copper, an active agent in many antifouling paints, and so have the potential to foul painted hulls (Piola and Johnston 2008).
Bugula neritina also has a positive economic impact, as a source of bryostatins, potential anticancer compounds (Davidson and Haygoood 1999). The extracted, purified compounds can be worth $380,000 per pound (Lovell et al. 2008).
Ecological Impacts on Chesapeake Native Species
Bugula neritina is currently rare in Chesapeake Bay, and probably has no detectable impacts on native biota. South of Cape Hatteras, and in subtropical-tropical waters worldwide, it is a major competitor in the fouling community (Sutherland and Karlson 1977; Winston 1982).
Ecological Impacts on Other Chesapeake Non-Native Species
Bugula neritina is currently rare in Chesapeake Bay, and probably has no detectable impacts on other introduced biota. South of Cape Hatteras, and in subtropical-tropical waters worldwide, it is a major competitor in the fouling community (Sutherland and Karlson 1977; Winston 1982).
References
Carlton, James T. (1979) History, biogeography, and ecology of the introduced marine and estuarine invertebrates of the Pacific Coast of North America, , Davis. Pp. 1-904Cohen, Andrew N.; Carlton, James T. (1995) Nonindigenous aquatic species in a United States estuary: a case study of the biological invasions of the San Francisco Bay and Delta, , Washington DC, Silver Spring MD.. Pp.
Coles, S. L.; DeFelice, R. C.; Eldredge, L. G.; Carlton, J. T. (1999b) Historical and recent introductions of non-indigenous marine species into Pearl Harbor, Oahu, Hawaiian Islands., Marine Biology 135: 147-158
Coues, Elliott; Yarrow, H.C. (1878) Notes on the natural history of Fort Macon, N.C. and vicinity (No. 5), Proceedings of the Academy of Natural Sciences of Philadelphia 30: 297-330
Cranfield, H.J.; Gordon, D.P.; Willan, R.C.; Marshall, B.A; Battershill, C.N.; Francis, M.P.; Nelson, W.A.; Glasby, C.J.; Read, G.B. (1998) Adventive marine species in New Zealand., , New Zealand. Pp.
Davidson, Seana K.: Haygood, Margo G. (1999) Identification of sibling species of the bryozoan Bugula neritina that produced different anticancer Bryostatins and harbor distinct strains of the bacterial symbiont "Candidatus endobugula sertula"., Biological Bulletin 196: 273-280
Gordon, D. P. (1986) The marine fauna of New Zealand: Bryozoa: Gymnolaemata (Ctenostomata and Cheilostomata Anasca) from the Western South Island continental shelf and slope., New Zealand Oceanographic Institute Memoir 95: 5-105
Gordon, Dennis P.; Mawatari, S.F. (1992) Atlas of marine-fouling bryozoa of New Zealand ports and harbors., Miscellaneous Publications of the New Zealand Oceanographic Institute 107: 1-52
Kaplan, Eugene H. (1988) A Field Gude to Southeastern and Caribbean Seashores, In: (Eds.) . , Boston. Pp.
Keough, M. J. ; Ross, J. (1999) Introduced fouling species in Port Phillip Bay., In: Hewitt, C. L.; Campbell; M.;Thresher, R.; Martin,(Eds.) Marine Biological Invasions of Port Phillip Bay, Victoria. , Hobart, Tasmania. Pp. 193-225
Keough, Michael J; Chernoff, Helen (1987) Dispersal and population variation in the Bryozoan Bugula neritina, Ecology 68: 199-210
Kitamura, Hitoshi; Hirayama, Kazutsugu (1984) Growth of the bryozoan Bugula neritina in the sea at various water temperatures., Bulletin of the Japanese Society of Scientific Fisheries 50: 1-5
Kitamura, Hitoshi; Hirayama, Kazutsugu (1985) The effect of low salinity on the early growth of a bryozoan Bugula neritina in the sea and laboratory., Bulletin of the Japanese Society of Scientific Fisheries 51: 1401-1405
Mackie, Joshua A.; Keough, Michael J.; Christidis, Les (2006) Invasion patterns inferred from cytochrome oxidase I sequences in three bryozoans, Bugula neritina, Watersipora subtorquata; and Watersipora arcuata., Marine Biology 149: 285-295
Marcus, Ernst (1937) Bryozoarios Marinhos Brasileiros, Boletim de faculdade de filosofia, Ciencias e Letras, Universidade de sao Paulo, Zoologia 1: 1-224
Maturo, Frank J. S., Jr. (1957) A study of the Bryozoa of Beaufort, North Carolina and vicinity, Journal of the Elisha Mitchell Scientific Society 72: 11-68
McGovern, Tamara M.; Hellberg, Michael E. (2003) Cryptic species, cryptic endosymbionts, and geographical variation in chemical defences in the Bryozoan Bugula neritina., Molecular Ecology 12: 1207-1215
Menon. N. R.; Nair, N. B. (1972) Indian species of the genus Bugula Oken., Proceedings of the Indian National Science Academy 38: 414-424
2003-2008 Introduced and cryptogenic species of the North Atlantic. http://massbay.mit.edu/exoticspecies/exoticmaps/introduced.html
Occhipinti Ambrogi, Anna; D'Hondt, Jean-Loup (1981) Distribution of bryozoans in brackish waters of Italy., In: Larwood, Gilbert P. and Nielsen, Claus(Eds.) Recent and Fossil Bryozoa.. , Fredensborg. Pp.
Orensanz, Jose Maria and 14 other authors (2002) No longer the pristine confines of the world ocean: a survey of exotic marine species in the southwestern Atlantic, Biological Invasions 4: 115-143
Osburn, Raymond C. (1932) Bryozoa from Chesapeake Bay, Ohio Journal of Science 32: 441-447
Osburn, Raymond C. (1940) Bryozoa of Porto Rico, N. Y. Academy of Sciences - Scientific Survey of Puerto Rico and the Virgin Islands 16: 321-486
Osburn, Raymond C. (1944) A survey of the Bryozoa of Chesapeake Bay, Chesapeake Biological Laboratory Publications 63: 1-55
Osburn, Raymond C. (1947) Bryozoa of the Allan Hancock Atlantic Expedition, Allan Hancock Atlantic Expedition Report 5: 1-66
Piola, Richard F.; Johnston, Emma L. (2006a) Differential resistance to extended copper exposure in four introduced bryozoans., Marine Ecology Progress Series 311: 103-114
Reise, K.; Gollasch, S.; Wolff, W.J. (1999) Introduced marine species of the North Sea coasts., Helgoländer Meeresuntersuchungen 52: 219-234
Ryland, J. S. (1960) The British species of Bugula (Polyzoa)., Proceedings of the Zoological Society of London 134: 65-105
Ryland, J. S. (1965) Catalogue of main marine fouling organisms- Bryozoa., None , Paris. Pp. None
Ryland, J. S. (1971) Bryozoa (Polyzoa) and marine fouling., In: Gareth Jones E.B; Eltringham, S. K.(Eds.) Marine Borers, fungi and fouling organisms of wood. , Paris. Pp. 137-154
Shier, Daniel (1964) Marine Bryozoa from Northwest Florida, Bulletin of Marine Science of the Gulf and Caribbean 14: 603-622
Sutherland, John P.; Karlson, Ronald H. (1977) Development and stability of the fouling community at Beaufort, North Carolina, Ecological Monographs 47: 425-446
Wendt, Dean E. (2000) Energies of larval swimming and metamorphosis in four species of Bugula (Bryozoa)., Biological Bulletin 198: 346-356
Winston, Judith E. (1977) Distribution and ecology of estuarine ectoprocts: a critical review., Chesapeake Science 18: 34-57
Winston, Judith E. (1982) Marine bryozoans (Ectoprocta) of the Indian River area (Florida)., Bulletin of the American Museum of Natural History 173: 99-176