Invasion History
First Non-native North American Tidal Record: 1936First Non-native West Coast Tidal Record: 1936
First Non-native East/Gulf Coast Tidal Record:
General Invasion History:
Heteromastus filiformis was described by Claparede in 1864 from Port-Vendres, on the Mediterranean coast of France, and later re-examined by Eisig, near Genoa, Italy in 1887. It was subsequently found on the East Coast of the US from Maine to North Carolina in the 1870s-80s (Hutchings and Rainier 1982). This worm has been reported in muddy-silty sediments on the coasts of every continent, except Antarctica. It spans Arctic and tropical waters, from the upper intertidal to the abyssal zone and from 5-40 PSU (Hartman 1947; Wolff 1973; Hartmann-Schroder 1980; Dean 2011; Weslawski et al. 2012; de Kluijver et al. 2014). Very likely 'H. filiformis’ represents a complex of cryptic species, similar to those found in its cosmopolitan relative, Capitella capitata (Eckelbarger and Grassle 1987).
However, the relatively recent collections and apparent spread of Heteromastus filiformis on the West Coast of North America, well after the onset of invertebrate collections, suggests that this polychaete has been introduced. It was first collected in San Francisco Bay in 1936, and found in harbors and bays from British Columbia (1st record 1962, Berkeley 1966) to Morro Bay (1st record 1960, Reish and Barnard 1967). Possible vectors include transport of larvae, or suspended adults in ballast water, and transport with plantings of Eastern Oysters (Crassostrea virginica). Occurrences in Prince William Sound, Alaska, may be the result of transport by shipping (1st record 1972, Hines et al. 2000), while deep water collections in the Bering Sea probably represent native Arctic cryptic species (US National Museum of Natural History 2015).
North American Invasion History:
Invasion History on the West Coast:
Heteromastus filiformis was first collected on the West Coast in San Francisco Bay, California (CA) in 1936 (Hartman 1947). It occurs at high densities in the central, South, and San Pablo Bays, and inland as far as Pittsburg, Suisun and Grizzly Bay (Carlton 1979; Nichols and Thompson 1985a; Nichols and Thompson 1985b; Peterson and Vayssieres 2010; California Academy of Sciences 2015). In 1962, this polychaete was collected at 24 m depth off Comox, Vancouver Island, in the Strait of Georgia (Berkeley 1966). Subsequent collections were made at Grays Harbor, Washington (1st record 1977, Albright and Smith 1977, cited by Carlton 1979); the Columbia River estuary (before 2002, Systsma et al. 2004); Coos Bay, Oregon (1st record 1970, Carlton 1989; Wonham and Carlton 2005); and Humboldt Bay, CA (2000, Boyd et al. 2002). More recently, it was collected in Southern California (Mission Bay and /Los Angeles-Long Beach Harbor in 2011 (California Department of Fish and Wildlife 2014).
However, specimens of 'Heteromastus filiformis' were collected in Alaskan offshore waters in the Bering Sea, near the Pribilof Islands at 135 m in 1976 (USNM 63505-63509, US National Museum of Natural History 2015), and in deep waters (524 m) off California (1988, USNM 123124, US National Museum of Natural History 2015). These deep water specimens presumably represent native Arctic or deep water members of the H. filiformis species complex. Specimens identified as H. filiformis occur in the Arctic Ocean, off Svalbard, down to 900m (Weslawski et al. 2012).
Invasion History Elsewhere in the World:
Heteromastus filiformis has been considered native or cryptogenic over much of its range. Introductions in ballast water, fouling, dry ballast, or with oysters are possible; however, given its small size, burrowing habitat, and paucity of distinguishing morphological features, it is likely that a number of cryptic species occupy its nominal range. Early records for Australia and New Zealand are 1969 in Port Philip Bay, Victoria (Hutchings and Rainier 1982) and 1976 in Hawke Bay, New Zealand (Knox and Fenwick 1981).
Description
Heteromastus filiformis is probably a complex of cryptic species belonging to the Capitellidae family. They lack protostomial or peristomial appendages, with branchiae absent or greatly reduced. The body has a distinct protostome, a thoracic version with a fixed number of segments, and a variable number of abdominal segments. Capitellids as a group are among the most abundant polychaetes in soft-bottom communities (Hartman 1947; Blake and Ruff 2007).
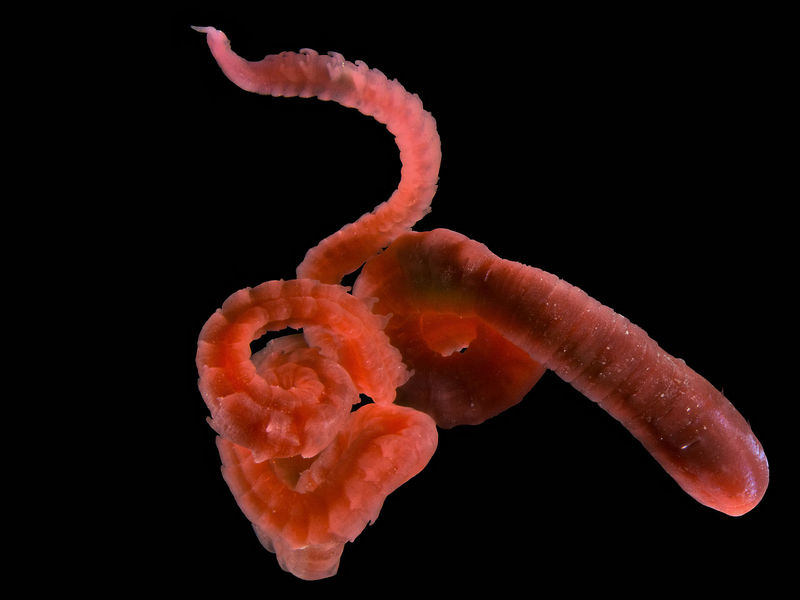
The body of H. filiformis is slender and fragile. The prostomium is small and conical, without eyespots or appendages. The thorax consists of 11 chaetigers and is slightly inflated anteriorly. Chaetigers 1-5 have capillary chaetae, while 6-11 have hooded hooks. The abdominal chaetae are all hooded hooks. From chaetigers 90-100, posteriorly, there are pairs of small, single, finger-like branchiae on each segment. The pygidium has a long, pointed terminal process. Specimens from the UK reach about 100 mm and have 140 segments (Hayward and Ryland 1990); specimens from the North Sea are about 180 mm with 150 segments; and specimens from the US East Coast are about 40 mm with 230 segments (Hutchings and Rainier 1982). Some of this size variation could be due to effects of preservation. The anterior region is dull red, while the posterior is reddish-yellow or greenish. Description based on: Hartman 1947, Hutchings and Rainier 1982, Hayward and Ryland 1990, Blake and Ruff 2007, and de Kluijver et al. 2014.
Limited comparative studies looking at the morphology, life history and genetics of H. filiformis populations have been conducted, to our knowledge. Hutchings and Rainier (1982) compared some populations and found no consistent morphological differences between specimens from Europe, the US East Coast, and Australia. Given the wide climate, habitat, and salinity range of records of H. filiformis, sibling speciation is likely. Molecular studies are needed to untangle the taxonomy, biogeography, and invasion history of the H. filiformis complex.
Morphology and development of H. filiformis larvae from Passamaquoddy Bay, New Brunswick are illustrated by Lacalli (1980).
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Annelida | |
| Class: | Polychaeta | |
| Order: | Capitellida | |
| Family: | Capitellidae | |
| Genus: | Heteromastus | |
| Species: | filiformis complex |
Synonyms
Notomastus filiformis (Verrill, 1873)
Areniella filiformis (Verrill, 1874)
Ancistria minima. (Webster, 1879)
Notomastus laevis (Webster, 1886)
Heteromastus filiformis (Eisig, 1887)
Potentially Misidentified Species
This capitlellid has multiple fine branchiae radiating out from its abdominal chaetifgers (Hartman 1947).
Mediomastus ambiseta
Juvenile Heteromastus filiformis have hooded hooks on chaetigers 4 and 5, and can be confused with Mediomastus spp. The hooded hooks are later replaced by capillary chaetae (Fredette 1982).
Mediomastus californiensis
Juvenile Heteromastus filiformis have hooded hox on chaetigers 4 and 5, and can be confused with Mediomastus spp. The hooded hooks are later replaced by capillary chaetae (Fredette 1982).
Ecology
General:
'Heteromastus filiformis' is a complex of cryptic species, found over a wide range of depth, climate, and salinity. Different populations are likely to vary in life history and environmental tolerances (Nygren 2014). Caution is advisable in generalizing information across regions. In European and Gulf of Maine populations, eggs are laid in gelatinous capsules on the sediment surface. Larvae are planktonic and planktotrophic (Lacalli 1980; Shaffer 1983). Larvae from Passamaquoddy Bay (Gulf of Maine) settled in ~17-18 days after hatching with ~11 setigers, at 400 μm length (Lacalli 1980). Adults in populations in North Inlet, South Carolina, lived for at least 2 years (Shaffer 1983).
'Heteromastus filiformis has been reported from Arctic, temperate, subtropical and tropical seas. West Coast populations are known from shallow estuaries and mudflats, and probably tolerate a wide temperature range (Nichols and Thompson 1985b; Hopkins 1986). In the Rhine Delta, Netherlands, 'H. filiformis' occurs at salinities as low as 5 PSU, but is much more common at salinities above 18 PSU (Wolff 1973). In the San Francisco estuary, this worm is common as far upstream as Grizzly Bay in dry years, at 5-10 PSU, but is more abundant in San Pablo Bay (Peterson and Vaysieres 2010). 'Heteromastus filiformis' is a head-down burrower, preferring muddy sediments, though it also occurs in fine and median sand (Wolff 1973; Quintana et al, 2007). It is a deposit-feeder, and can burrow up to 15 cm beneath the sediment surface. The worm ventilates its burrow, and the oxygenated water in the burrow allows the worms to penetrate for some distance into anoxic sediments. Feeding is believed to be non-selective, on microalgae, detritus, and other organic particles (Wolff 1973). Fecal pellets are deposited on the sediment surface (Quintana et al. 2007). 'Heteromastus filiformis' often reaches high abundances, and is potentially an important food item for benthic invertebrates and fishes. However, caging experiments, in North Inlet, South Carolina, suggested that excluding large predators had no effect on this species density (Shaffer 1982).
Food:
Detritus
Trophic Status:
Deposit Feeder
DepFedHabitats
| General Habitat | Grass Bed | None |
| General Habitat | Unstructured Bottom | None |
| General Habitat | Salt-brackish marsh | None |
| Salinity Range | Mesohaline | 5-18 PSU |
| Salinity Range | Polyhaline | 18-30 PSU |
| Salinity Range | Euhaline | 30-40 PSU |
| Tidal Range | Subtidal | None |
| Tidal Range | Low Intertidal | None |
| Vertical Habitat | Endobenthic | None |
Tolerances and Life History Parameters
| Minimum Salinity (‰) | 5 | Field, But uncommon, below 18 PSU, Netherlands (Wolff 1973) |
| Maximum Salinity (‰) | 40 | average Red Sea salinity (Wehe and Fiege 2002) |
| Maximum Duration | 18 | Larvae from Passamaquoddy Bay (Gulf of Maine) settled in ~17-18 days after hatching (Lacalli 190). Wolff (1973) gave an estimated planktonic larval duration of 10 days for worms from the Netherlands, based on filed data. |
| Maximum Length (mm) | 180 | Hutchings and Rainer (1982). The pygidium has a long, pointed terminal process. Specimens from the UK reach about 100 mm and have 140 segments (Hayward and Ryland 1990); specimens from the North Sea are about 180 mm with 150 segments; and specimens from the US East Coast are about 40 mm with 230 segments (Hutchings and Rainier 1982). Some of this size variation could be due to effects of preservation. |
| Broad Temperature Range | None | Cold temperate-Tropical |
| Broad Salinity Range | None | Oligohaline-Euhaline |
General Impacts
Capitellid polychaetes, including Heteromastus filiformis, are an important component of infaunal communities, burrowing and reworking sediments, and increasing the oxygen penetration (Wolff 1973; Quintna et al. 2007). They are a potentially important food source for benthic predators. However, the specific impacts of H. filiformis are difficult to determine, since it coexists with many other native and cryptogenic capitellid polychaetes (Hartman 1947; Blake and Ruff 2007).Regional Distribution Map
Non-native
Native
Cryptogenic
Failed
| Bioregion | Region Name | Year | Invasion Status | Population Status |
|---|---|---|---|---|
| NEP-VI | Pt. Conception to Southern Baja California | 2011 | Non-native | Established |
| P030 | Mission Bay | 2011 | Non-native | Established |
| P050 | San Pedro Bay | 2011 | Non-native | Established |
| P130 | Humboldt Bay | 2000 | Non-native | Established |
| P080 | Monterey Bay | 1977 | Non-native | Established |
| NEP-IV | Puget Sound to Northern California | 1970 | Non-native | Established |
| P095 | _CDA_P095 (Tomales-Drakes Bay) | 1969 | Non-native | Established |
| P070 | Morro Bay | 1960 | Non-native | Established |
| P093 | _CDA_P093 (San Pablo Bay) | 1936 | Non-native | Established |
| NEP-V | Northern California to Mid Channel Islands | 1936 | Non-native | Established |
| P090 | San Francisco Bay | 1936 | Non-native | Established |
Occurrence Map
| OCC_ID | Author | Year | Date | Locality | Status | Latitude | Longitude |
|---|---|---|---|---|---|---|---|
| 755700 | Introduced Species Study | 2003 | 2003-08-27 | Corinthian Marina 2 | Non-native | 37.8720 | -122.4556 |
| 755701 | Introduced Species Study | 2003 | 2003-08-27 | Loch Lomond 2 | Non-native | 37.9717 | -122.4811 |
| 755702 | Introduced Species Study | 2005 | 2005-06-07 | Oakland Inner Harbor - Small marinas | Non-native | 37.7847 | -122.2669 |
| 755703 | Introduced Species Study | 2005 | 2005-06-07 | Port of Oakland Office | Non-native | 37.7954 | -122.2804 |
| 755704 | Introduced Species Study | 2005 | 2005-06-08 | Oakland Outer Harbor | Non-native | 37.8217 | -122.3145 |
| 755705 | Introduced Species Study | 2005 | 2005-06-08 | Sea Plane Lagoon | Non-native | 37.7761 | -122.2998 |
| 755706 | Introduced Species Study | 2005 | 2005-06-09 | Paradise Area | Non-native | 37.9062 | -122.4768 |
| 755707 | Introduced Species Study | 2005 | 2005-06-09 | Richardson Bay | Non-native | 37.8588 | -122.4798 |
| 755708 | Introduced Species Study | 2005 | 2005-06-10 | Hayward Landing | Non-native | 37.6447 | -122.1543 |
| 755709 | Introduced Species Study | 2005 | 2005-06-10 | Toll Plaza | Non-native | 37.8266 | -122.3166 |
| 755710 | Introduced Species Study | 2005 | 2005-08-25 | Ferry Terminal Pier | Non-native | 37.7945 | -122.3917 |
| 755711 | Introduced Species Study | 2005 | 2005-09-07 | Redwood Creek - Marina | Non-native | 37.5021 | -122.2130 |
| 755712 | Introduced Species Study | 2005 | 2005-09-07 | Redwood Creek - Shipping | Non-native | 37.5120 | -122.2109 |
| 755713 | Introduced Species Study | 2005 | 2005-09-09 | Coyote Point Marina | Non-native | 37.5905 | -122.3177 |
| 755714 | Introduced Species Study | 2005 | 2005-09-09 | San Mateo Bridge | Non-native | 37.5806 | -122.2543 |
| 755715 | Introduced Species Study | 2005 | 2005-09-09 | Sierra Point Marina | Non-native | 37.6740 | -122.3792 |
| 755716 | Introduced Species Study | 2005 | 2005-10-06 | Santa Fe Channel - Back | Non-native | 37.9207 | -122.3684 |
| 755717 | Introduced Species Study | 2005 | 2005-10-19 | Hercules Wharf | Non-native | 38.0231 | -122.2928 |
| 755718 | Introduced Species Study | 2005 | 2005-10-19 | Mare Island Strait - Marina | Non-native | 38.1051 | -122.2667 |
| 755719 | Introduced Species Study | 2005 | 2005-10-20 | Port Sonoma/Petaluma R. | Non-native | 38.1157 | -122.5026 |
| 755720 | Introduced Species Study | 2005 | 2005-10-20 | San Pablo Bay Pumphouse | Non-native | 38.0446 | -122.4326 |
| 755721 | Introduced Species Study | 2005 | 2005-10-21 | Ayala Cove | Non-native | 37.8680 | -122.4350 |
| 755722 | Introduced Species Study | 2010 | 2010-06-13 | Hayward Landing | Non-native | 37.6447 | -122.1543 |
| 755723 | Introduced Species Study | 2010 | 2010-07-01 | Corinthian Marina | Non-native | 37.8726 | -122.4563 |
| 755724 | Introduced Species Study | 2011 | 2011-04-20 | Slip D-50 | Non-native | 33.7165 | -118.2801 |
| 755725 | Introduced Species Study | 2011 | 2011-05-04 | Seaforth | Non-native | 32.7621 | -117.2365 |
| 758921 | Hartman 1947 | 1936 | San Francisco Bay | Non-native | 37.8494 | -122.3681 | |
| 758922 | Hartman 1954 | 1954 | San Pablo Channel | Non-native | 37.9698 | -122.4385 | |
| 758923 | Reish and Barnard 1967 | 1960 | 1960-08-25 | Morro Bay | Non-native | 35.3500 | -120.8500 |
| 758924 | Jones 1961 | 1955 | Point Richmond, Station A | Non-native | 37.9211 | -122.3881 | |
| 758925 | Jones 1961 | 1955 | Point Richmond, Station B | Non-native | 37.9114 | -122.3889 | |
| 758926 | Jones 1961 | 1955 | Point Richmond, Station C | Non-native | 37.9036 | -122.3853 | |
| 758927 | Light 1969 | 1967 | Bolinas Lagoon | Non-native | 37.9189 | -122.6816 | |
| 758928 | Chapman and Dorman 1975 | 1972 | 1972-03-19 | Tubbs Island, NE side | Non-native | 38.1256 | -122.4367 |
| 758929 | Nybakken et al. 1977 | 1974 | Station N-2, offshore of Moss Landing Harbor | Non-native | 36.8144 | -121.7992 | |
| 758930 | Nybakken et al. 1977 | 1975 | 1975-09-16 | Station N-2, offshore of Moss Landing Harbor | Non-native | 36.8144 | -121.7992 |
| 758931 | Nybakken et al. 1977 | 1975 | 1975-04-02 | Station N-4, offshore of Moss Landing Harbor | Non-native | 36.8099 | -121.8094 |
| 758932 | Nybakken et al. 1977 | 1976 | 1976-06-15 | Beach in front of original Moss Landing Marine Laboratories site (Station 11-1B) | Non-native | 36.8002 | -121.7897 |
| 758933 | Nichols and Thompson 1985a | 1985 | San Pablo Bay | Non-native | 38.0600 | -122.3900 | |
| 758934 | Nichols and Thompson 1985a, 1985b | 1985 | Sand Point, Palo Alto | Non-native | 37.4630 | -122.1011 | |
| 758935 | Boyd et al. 2002 (Humboldt Bay Report) | 2002 | Bracut | Non-native | 40.8313 | -124.0845 | |
| 758936 | Boyd et al. 2002 (Humboldt Bay Report) | 2002 | Eureka Channel HB, St. 21 | Non-native | 40.8063 | -124.1666 | |
| 758937 | Boyd et al. 2002 (Humboldt Bay Report) | 2002 | Hilfiker Road | Non-native | 40.7720 | -124.1960 | |
| 758938 | Boyd et al. 2002 (Humboldt Bay Report) | 2002 | Jacoby Creek | Non-native | 40.8435 | -124.0838 | |
| 758939 | Boyd et al. 2002 (Humboldt Bay Report) | 2002 | Klopp Lake | Non-native | 40.8553 | -124.0919 | |
| 758940 | Boyd et al. 2002 (Humboldt Bay Report) | 2002 | Mad River Slough - Samoa Blvd. Bridge | Non-native | 40.8652 | -124.1505 | |
| 758941 | Cohen et al. 2005 (SF Bay Area RAS) | 2004 | 2004-05-28 | Rodeo Marina, San Pablo Bay | Non-native | 38.0391 | -122.2711 |
| 758942 | Heiman and Micheli 2010; Hobbs et al. 2015 | 2002 | Elkhorn Slough (Upper) | Non-native | 36.8547 | -121.7600 |
References
Berkeley, C. (1966) Records of some Polychaeta new to British Columbia new to British Columbia and of extensions in distribution of some others, Canadian Journal of Zoology 44: 839-848Blake, James A.; Ruff, R. Eugene (2007) The Light and Smith Manual: Intertidal invertebrates from Central California to Oregon (4th edition), University of California, Berkeley CA. Pp. 309-410
Boyd, Milton J.; Mulligan, Tim J; Shaughnessy, Frank J. (2002) <missing title>, California Department of Fish and Game, Sacramento. Pp. 1-118
California Academy of Sciences 2005-2015 Invertebrate Zoology Collection Database. <missing URL>
California Department of Fish and Wildlife (2014) Introduced Aquatic Species in California Bays and Harbors, 2011 Survey, California Department of Fish and Wildlife, Sacramento CA. Pp. 1-36
Carlton, James T. (1979) History, biogeography, and ecology of the introduced marine and estuarine invertebrates of the Pacific Coast of North America., Ph.D. dissertation, University of California, Davis. Pp. 1-904
Carlton, James T. (1989) <missing title>, <missing publisher>, <missing place>. Pp. <missing location>
Çinar, Melih Ertan; Dagli, Ertan (2013) Polychaetes (Annelida: Polychaeta) from the Aegean and Levantine coasts of Turkey, with descriptions of two new species, Journal of Natural History 47(13-14): 911-947
Çinar, Melih Ertan; Dagli, Ertan; Açik, Sermin (2011) Annelids (Polychaeta and Oligochaeta) from the Sea of Marmara, with descriptions of five new species, Journal of Natural History 45(33-34): 2105-2143
Cohen, Andrew N. and 10 authors (2005) <missing title>, San Francisco Estuary Institute, Oakland CA. Pp. <missing location>
Cohen, Andrew N.; Carlton, James T. (1995) Nonindigenous aquatic species in a United States estuary: a case study of the biological invasions of the San Francisco Bay and Delta, U.S. Fish and Wildlife Service and National Sea Grant College Program (Connecticut Sea Grant), Washington DC, Silver Spring MD.. Pp. <missing location>
Collado-Vides, L. (2002) Morphological plasticity of Caulerpa prolifera (Caulerpales, Chlorophyta) in relation to growth form in a coral reef lagoon, Botanica Marina 45: 123-129
de Kluijver, Mario; Ingalsuo, Sarita.; van Nieuwenhuijzen, Andre; Veldhuijzen-Van Zanten, Huub 2014 Marine Species Identification Portal: Macrobenthos of the North Sea - Polychaeta. <missing URL>
Dean, Harlan K. (2011) Capitellidae (Annelida: Polychaeta) from the Pacific Coast of Costa Rica, Revista Biologia Tropical 49(2): 69-84
Dewarumez, M.; Davoult, D.; Glaçon, R. (1992) [New records of macrobenthic species on the French coasts of the eastern English Channel and the North Sea. 1. Annelids.], Cahiers de Biologie Marine 33: 83-93
Fredette, Thomas J. (1982) Evidence of ontogeneitc setal changes in Heteromastus filiformis (Polycaheta: Capitellidae), Proceedings of the Biological Society of Washington 95(`): 194-197
Hartman, Olga (1947) Polychaetous annelids. Part VII. Capitellidae, Allan Hancock Pacific Expeditions 10: 391-480
Hartman, Olga (1969) <missing title>, Allan Hancock Foundation, Los Angeles. Pp. <missing location>
Hartmann-Schroder, Gesa (1980) Amsterdam Expeditions to the West Indian Islands, Report 9. The polychaetes of the Amsterdam Expeditions to the West Indian Islands., Bijdragen tot de Dierkunde 50(2): 387-401
Hines, Anson H.; Ruiz, Gregory M. (2000) Biological invasions of cold-water coastal ecosystems: ballast-mediated introductions in Port Valdez/Prince William Sound (Final Report), In: (Eds.) . , Valdez, Alaska. Pp. <missing location>
Hines, Anson H.; Ruiz, Gregory M.; Chapman, John; Hansen, Gayl; Carlton, James T.; Foster, Nora; Feder, Howard M. (1998) <missing title>, Regional Citizens Advisory Council of Prince William Sound, Valdez, Alaska.. Pp. <missing location>
Hopkins, Dale R. (1986) Atlas of the distributions and abundances of common benthic species in San Francisco Bay, California, US Geological Survey Water Resources Investigations Report 86-4003: 1-16+ 25+228
Huang, Zongguo (Ed.), Junda Lin (Translator) (2001) Marine Species and Their Distributions in China's Seas, Krieger, Malabar, FL. Pp. <missing location>
Hutchings, P. A.; Rainier, S. (1982) Designation of a neotype of Capitella filiformis Claparède, 1864, type species of the genus Heteromastus (Polychaeta: Capitellidae), Records of the Australian Museum 34(4): 373-380
Knox, G. A.; Fenwick, Graham D. (1981) Zonation of inshore benthos off a sewage outfall in Hawke Bay, New Zealand, New Zealand Journal of Marine and Freshwater Research 15: 417-435
Morgan, Vanessa H.; Sytsma, Mark D. (2013) Potential ocean dispersal of cordgrass (Spartina sp.) from core infestations, Invasive Plant Science and Management 6: 250-259
Nichols, Frederic H.; Thompson, Janet K. (1985a) Time scales of change in the San Francisco Bay benthos., Hydrobiologia 129: 121-138
Nichols, Frederic H.; Thompson, Janet K. (1985b) Persistence of an introduced mudflat community in South San Francisco Bay, California, Marine Ecology Progress Series 24: 83-97.
Peterson, Heather A.; Vayssieres, Marc (2010) Benthic assemblage variability in the upper San Francisco estuary: A 27-year retrospective, San Francisco Estuary and Watershed Science <missing volume>: published online
Quintana, Cintia Organo; Tang, Min; Kristensen, Erik (2007) Simultaneous study of particle reworking, irrigation transport and reaction rates in sediment bioturbated by the polychaetes Heteromastus and Marenzelleria, Journal of Experimental Marine Biology and Ecology 352: 392-406
Reish, Donald J.; Barnard, J. Laurens (1967) The benthic Polychaeta and Amphipoda of Morro Bay, California, Proceedings of the United States National Museum 120(3565): 1-26
Riggs, Sharon R. (2011) <missing title>, Padilla Bay NERR, Padilla Bay WA. Pp. 5
Sahin, Güley Kurt; Çinar, Melih Ertan (2012) A check-list of polychaete species (Annelida: Polychaeta) from the Black Sea, Journal of the Black Sea/Mediterranean Environment 18(1): 10-48
Shaffer, P. I. (1983) Population ecology of Heteromastus filiformis (Polychaeta: Capitellidae), Journal of Sea Research 17(1): 106-125
Sytsma, Mark D.; Cordell, Jeffrey R.; Chapman, John W.; Draheim, Robyn, C. (2004) <missing title>, Center for Lakes and Reservoirs, Portland State University, Portland OR. Pp. <missing location>
U.S. National Museum of Natural History 2002-2021 Invertebrate Zoology Collections Database. http://collections.nmnh.si.edu/search/iz/
Wakeman, Kevin C.; Leander, Brian S. (2013) Identity of environmental DNA sequences using descriptions of four novel marine gregarine parasites, Polyplicarium n. gen. (Apicomplexa), from capitellid polychaetes, Marine Biodiversity Published online: <missing location>
Wasson, Kerstin; Zabin, C. J.; Bedinger, L.; Diaz, M. C.; Pearse J. S. (2001) Biological invasions of estuaries without international shipping: the importance of intraregional transport, Biological Conservation 102: 143-153
Wehe, Thomas; Fiege, Dieter (2002) Annotated checklist of the polychaete species of the seas surrounding the Arabian Peninsula: Red Sea, Gulf of Aden, Arabian Sea, Gulf of Oman, Arabian Gulf., Fauna of Arabia 19: 7-238
Weslawski, Jan Marcin; Maria Wlodarska-Kowalczuk, Maria; Kedra, Monika; Joanna Legeynska; Kotwiki, Lech (2012) Eight species that rule today’s European Arctic nfjord benthos, Polish Polar Research 33(3): 225-238
Wolff, W. J. (1973) The estuary as a habitat: An analysis of data on the soft-bottom macrofauna of the estuarine area of the rivers Rhine, Meuse, and Scheldt, Zoologische Verhandelingen 126: 4-242
Wonham, Marjorie J.; Carlton, James T. (2005) Trends in marine biological invasions at local and regional scales: the Northeast Pacific Ocean as a model system, Biological Invasions 7: 369-392