Invasion History
First Non-native North American Tidal Record:First Non-native West Coast Tidal Record:
First Non-native East/Gulf Coast Tidal Record:
General Invasion History:
None
North American Invasion History:
Description
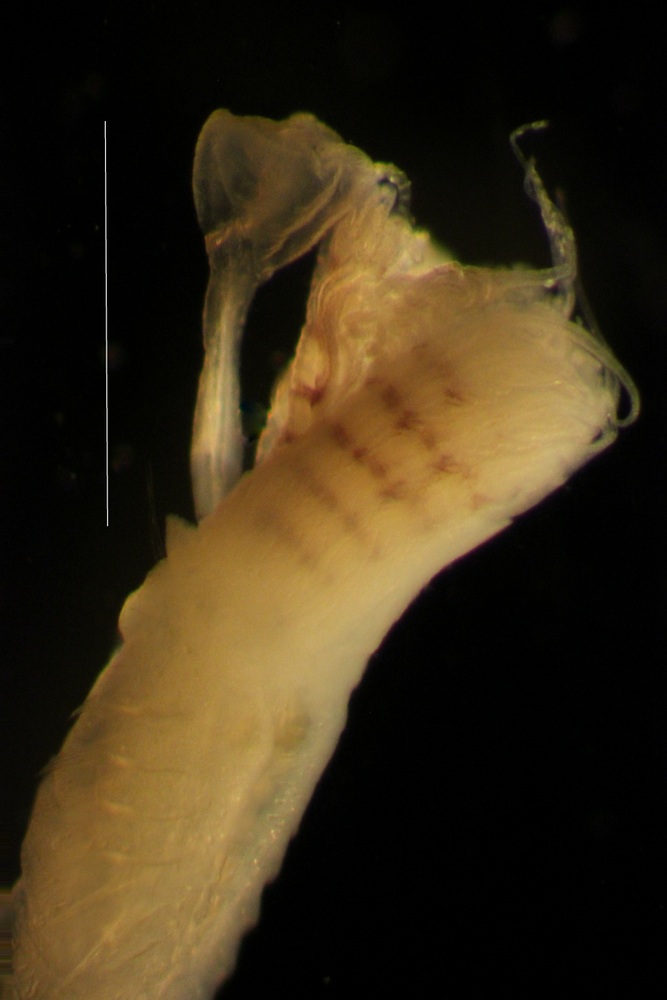
Ficopomatus miamiensis secretes a calcareous tube, as do other serpulid polychaetes. Serpulids have a feathery crown of modified prostomial palps, called radioles (the prostomium is the first segment, projecting above the mouth). The radioles can be folded and withdrawn into the tube. One of the radioles is modified to form an operculum, which acts as a plug when the animal contracts.
The peristomium (segment behind the mouth) is folded back to form a collar, which bears uniramous parapodia, with a distinctive set of collar chaetae, with spines or serrations. The collar is the first of seven thoracic chaeta-bearing segments (setigers). The subsequent segments have biramous parapodia. The dorsal branch of the parapodium is called the notopodium; the ventral is the neuropodium. Chaetae in the two branches and along the body can vary greatly in their morphology, which can be critical in the taxonomy of serpulids (Description from: Barnes 1983; ten Hove and Weerdenburg 1978; Blake and Ruff, in Carlton 2007).
The tube of F. miamiensis is shining white, and circular in cross-section, and lacks rings or keels. Solitary and juvenile tubes may have faint longitudinal keels.. lThe branchial (gill) radioles arise from paired lobes. There are about 7 (5-9) on the left and 8 (6-10) on the right, forming two semicircles on either side of the mouth. The feathery filaments of the branchiae (pinnulae) are shorter towards the ends of the radioles. The operculum is shaped like an elongated funnel, with a slightly concave, smootjh, distal surface, lacking spijne. The gills and operculum account for about 1/6 of the length of the worm. The peduncle of the operculum is D-shaped in cross-section. The operculam is fig-shaped and convex, lacking spines.The collar (first thoracic segment) is high and not lobed, with a smooth edge. The thorax consists of 7 segments. There are two kinds of collar chaetae, thicker coarsely serrated chaetae, and hair-like (limbate). The subsequent thoracic segments bear short, rasplike chaetae, called uncinae. and limbate chaetae. The abdomen has about 40 segments (23-58 n=7, ten Hove and Weerdenburg 1978). The overall length of the worm reaches about 7 (2.5-11mm). (ten Hove and Weerdenburg 1978;
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Annelida | |
| Class: | Polychaeta | |
| Subclass: | Palpata | |
| Order: | Canalipalpata | |
| Suborder: | Sabellida | |
| Family: | Serpulidae | |
| SubFamily: | Serpulinae | |
| Genus: | Ficopomatus | |
| Species: | miamiensis |
Synonyms
Potentially Misidentified Species
None
Ficopomatus uschakovi
None
Ecology
General:
The serpulid polychaetes og the genus Ficopomatus feed by extending its feathery gills to trap plankton in the water column. The plankton is then transported by cilia to the mouth. In most populations of F.enigmaticu,, the two sexes are separate (Tunisia- Vuillemin 1965; Argentina- Obenat and Pezzani 1994), but in at least one population (Thames estuary, England), a small percentage of hermaphroditic individuals were seen (Dixon probably 1981). Details of spawning are not known,, but eggs and sperm are released into the water column, and the larvae are planktotrophic.
Ficopomatus miamiensis tolerates subtropical -to tropcial temperatures and brackish to hyperhaline salinities (2.5-43 PSU). . Habitiats include creeks an d canals, shrimp ponds, mangroves, and the walls of the Panama Canal (Ten Hove, and Weerdenburg, 1978; Salgado-Barragan,2004)
At the northern limits of its range in North America and Europe, its establishment seems to be favored by thermal effluents (ten Hove 1974: Turner and Hoagland 1980; Jensen and Knudsen 2005). A temperature of 18C is apparently required for spawning (Dixon 1981). This tubeworm tolerates salinities ranging from 1 to 55 PSU, but seems especially abundant in brackish waters (ten Hove and Weerdenberg 1978), possibly because of a lack of competition. It settles and secretes a calcareous tube on surfaces such as rocks, shells, plant leaves, pilings and pontoon floats. While F. enigmaticus is widespread, reef development does not always occur. Shallow estuaries and lagoons with long retention times appear especially prone to reef formation. The reefs begin as clumps of intertwined tubes, attached to sticks, stone, shells etc., and grow over many generations of settlement. Reefs can reach 0.5 m in thickness and 7 m across (Bianchi and Morri 1996; Schwindt et al. 2001).
Food:
Phytoplankton, detritus
Trophic Status:
Suspension Feeder
SusFedHabitats
| General Habitat | Coarse Woody Debris | None |
| General Habitat | Salt-brackish marsh | None |
| General Habitat | Oyster Reef | None |
| General Habitat | Marinas & Docks | None |
| General Habitat | Rocky | None |
| General Habitat | Mangroves | None |
| General Habitat | Canals | None |
| Tidal Range | Subtidal | None |
| Vertical Habitat | Epibenthic | None |
Tolerances and Life History Parameters
| Minimum Salinity (‰) | 2.5 | Field, Lake Pontchartrain (ten Hove and Weerdenburg 1978) |
| Maximum Salinity (‰) | 46 | Field, Urias estuary, Mexico (Salgado-Barragan et al. 2014) |
| Maximum Length (mm) | 10 | None |
| Broad Temperature Range | None | Subtropical-Tropical |
| Broad Salinity Range | None | Oligohaline-Hyperhaline |
General Impacts
Regional Distribution Map
| Bioregion | Region Name | Year | Invasion Status | Population Status | Vectors |
|---|---|---|---|---|---|
| SEP-H | None | 1972 | Non-native | Established | |
| CAR-I | Northern Yucatan, Gulf of Mexico, Florida Straits, to Middle Eastern Florida | 1934 | Native | Established | |
| CAR-II | None | 0 | Native | Established | |
| CAR-IV | None | 0 | Native | Established | |
| CAR-III | None | 1970 | Native | Established | |
| NEP-VIII | None | 1993 | Non-native | Established | |
| NEP-VII | None | 2012 | Non-native | Established | |
| PAN_PAC | Panama Pacific Coast | 1972 | Non-native | Established |
Occurrence Map
| OCC_ID | Author | Year | Date | Locality | Status | Latitude | Longitude |
|---|
References
Bastida-Zavala, J. Rolando; McCann, Linda D.; Keppel; Erica; Ruiz, Gregory M. (2017) The fouling serpulids (Polychaeta: Serpulidae) from United States coastal waters: an overview, European Journal of Taxonomy 344: 1-76Bastida-Zavala, J. Rolando. (2008) Serpulids (Annelida: Polychaeta) from the Eastern Pacific, including a brief mention of Hawaiian serpulids., Zootaxa 1722: 1-61
Bastida-Zavala, Jose Rolando; Salazar-Vallejo, Sergio (2000) Serpulidos (Polychaeta: Serpulidae) del Caribe norocidental: Hydroides Y Serpula, Revista de Biologia Tropical 48(4): 841-858
Bastida-Zavala, Rolando; de León-González, Jesús Ángel; Carballo Cenizo, José Luis; Moreno-Dávila, Betzabé (2014) [Aquatic Invasive Species in Mexico], Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, <missing place>. Pp. 317-336
Blakeslee, April M. H.; Miller, A. Whitman; Ruiz, Gregory M.; Kerstin Johannesson · Carl André ·Johannsson, Kerstin;· André, Carl; Panova, Marina (2021) Population structure and phylogeography of two North Atlantic Littorina species with contrasting larval development, Marine Biology 168: <missing location>
Hernández, María Ana Tovar; de León-González, Jesús Angel; Kupriyanova. Elena (2022) New records of invasive tubeworms (Ficopomatus, Serpulidae) in Mexico, Journal of the Marine Biological Association of the United Kingdom 102: 553-564
https://doi.org/10.1017/ S0025315422000790
Low-Pfeng, Antonio; Recagno, Edward M. Peters (2012) <missing title>, Geomare, A. C., INESEMARNAT, Mexico. Pp. 236
Salgado-Barragan, Jose; Mendez, Nutria; Toledano-Granados, Arturo (2004) Ficopomatus miamiensis (Polychaeta: Serpulidae) and Styela canopus (Ascidiacea: Styelidae), non-native species in Urias estuary, SE Gulf of California, Mexico., Cahiers de Biologie Marine 45: 167-173
Ten Hove, H. A.; Weerdenburg, J. C. A. (1978) A generic revision of the brackish-water serpulid Ficopomatus Southern 1921 (Polychaeta : Serpulinae) including Mercierella Fauvel 1923, Sphaeropomatus Treadwell 1934, Mercierellopsis Rioja 1945 and Neopomatus Pillai 196, Biological Bulletin 154: 96-120
Tovar-Hernández, M. A.; Villalobos-Guerrero, T. F.; Yáñez-Rivera, B., Aguilar-Camacho, J. M.; Ramírez-Santana, I. D. (2012) [Guide to exotic aquatic invertebrates in Sinaloa] , Geomare, A. C., USFWS, INE-SEMARNAT, Mazatlán, México. Pp. 41
Tovar-Hernandez, Maria Ana; Mendez, Nuria; Villalobos-Guerrero, Tulio Fabio (2009) Fouling polychaete worms from the southern Gulf of California: Sabellidae and Serpulidae, Systematics and Biodiversity 7(3): 319-336
U.S. National Museum of Natural History 2002-2021 Invertebrate Zoology Collections Database. http://collections.nmnh.si.edu/search/iz/