Invasion History
First Non-native North American Tidal Record: 2006First Non-native West Coast Tidal Record: 2010
First Non-native East/Gulf Coast Tidal Record: 2006
General Invasion History:
Turritopsis dohrnii was first described from Naples, Italy, and is a common hydroid on rocky substrates in the Western Mediterranean and Adriatic. However, it is less well-known in this region as a medusa (Schuchert 2004). In a survey of hydroids in Lebanon, it was collected only once, in a harbor (Morri et al. 2009). Most reports of T. nutricula in the Mediterranean probably refer to T. dohrnii, but the presence of T. nutricula or other species, cannot be excluded (Schuchert 2004). Miglietta et al. (2006) and Miglietta and Lessios (2009) conducted a genetic survey of Turritopsis spp., and found T. dohrnii in the Indian River Lagoon, Florida; both the Caribbean and Pacific coasts of Panama; and Japan. They also found a high degree of genetic similarity among populations. They concluded that this disjunct distribution could not be explained by long-distance dispersal of medusae in ballast water, or polyps on ships’ hulls. An unusual feature of T. dohrnii's life cycle is the ability of medusae to transform into a cyst, which can then regenerate into a polyp, making the organism potentially immortal (Miglietta and Lessios 2009).
North American Invasion History:
Invasion History on the West Coast:
In 2019 two samples of colonies from Newport Beach, California were genetically identified as T. dohrnii (Miglietta et al. 2019).
Invasion History on the East Coast:
Medusae and polyps of Turritopsis dohrnii were collected at Fort Pierce, Florida in 2006, in the Indian River Lagoon, and identified using 16S mitochondrial DNA (Miglietta and Lessios 2009).
Invasion History Elsewhere in the World:
Miglietta and Lessios (2009) surveyed populations of Turritopsis spp. at many locations in the Atlantic and Pacific in 2002-2009. On the Caribbean coast of Panama, medusae of T. dohrnii were found at Galeta Island, near the Panama Canal entrance, this hydrozoan was also found in many samples in Bocas del Toro, about 200 km west of the Canal. On the Pacific side of Panama, medusae of T. dohrnii were found in Panama Bay, and were one of the more abundant hydromedusae present (Miglietta and Lessios 2009). In Japan, polyps of this species were found in 2002 in 'southern Japan' and in 2003 on Okinawa (Miglietta et al. 2006; Miglietta and Lessios 2009).
Description
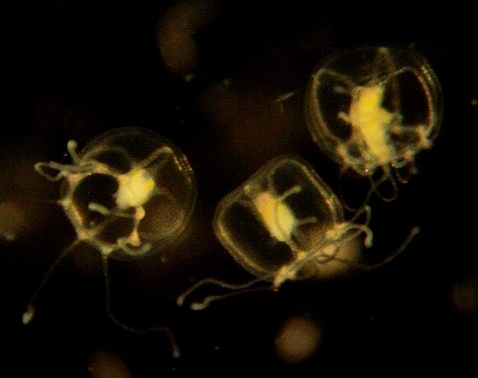
Turritopsis dohrnii is a hydrozoan with both a polyp and medusa stage. The hydroid colonies vary from sparingly branched colonies a few mm high to much-branched, multiple-stemmed colonies up to 35 mm high. The hydranths branch off at an acute angle from the hydrocaulus (stem), and then curve away. The stem and branches are covered by a firm perisarc, often encrusted with detritus and algae. The hydranths are spindle-shaped and have 12-20 filiform (thread-like) tentacles, spread over most of their length, and end in a conical hypostome. The pear-shaped medusa buds arise singly on short stems from below the hydranths. The hydranths can be colorless or pinkish.
Newly released medusae have 8-12 tentacles, and a conical manubrium, with four fluorescent-yellow pads between the roots of the interradial canals. The adult medusae are up to 2.7 mm in height and 3.3 mm in diameter, with 14-32 tentacles. Numbers of tentacles in adult medusae vary regionally with: 8 in Bocas del Toro, Panama; 12-24 in Italy; and 14-24 in Japan, probably in part due to environmental influences. The tentacles sometimes have terminal swellings. The manubrium is cross-shaped in cross section, ending in four lips, and reaches to the bell margin. It is red, while the gonads are brownish, with four rust-colored dots, between the radial canals. (Description from: Bouillon et al. 2004; Schuchert 2004)
Medusae of the genus Turritopsis have been widely identified as T. nutricula McCrady, 1857 (described from Charleston, South Carolina) around the world. Turritopsis dohrnii is often treated as a synonym of T. nutricula; however, different populations have been shown to have different morphologies and life histories, both in the medusa and polyp stage (Schuchert 2004). A genetic study of Turritopsis identified seven species from the Atlantic and Pacific, four of which are unnamed (Miglietta et al. 2006). Turritopsis dohrnii, described from the Mediterranean, has a disjunct distribution and genetic similarity indicative of human-mediated introduction (Miglietta and Lessios 2009). Additional cryptic species of Turritopsis may exist, and other 'silent invasions' are possible. Identification of the cryptic species of Turritopsis is only possible by molecular means (Miglietta and Lessios 2009).
Turritopsis dohrnii has been christened 'Immortal Jellyfish' in the popular press, because starved medusae can settle to become an undifferentiated cyst, which grows into a new polyp, restartimg the life cycle. This process of transdifferentiation makes each individual organism potentially immortal (Miglietta et al. 2009).
Taxonomy
Taxonomic Tree
| Kingdom: | Animalia | |
| Phylum: | Cnidaria | |
| Class: | Hydrozoa | |
| Subclass: | Hydroidolina | |
| Order: | Anthoathecatae | |
| Suborder: | Filifera | |
| Family: | Oceaniidae | |
| Genus: | Turritopsis | |
| Species: | dohrnii |
Synonyms
Tubiclava fruticosa (Riedl, 1959)
Potentially Misidentified Species
NW Atlantic (Woods Hole, Miglietta et al. 2009)
Turritopsis rubra
W Pacific (Japan, New Zealand, Miglietta et al. 2009)
Turritopsis sp. 1
W Atlantic (Panama, Miglietta et al. 2009)
Turritopsis sp. 2
NW Pacific (Japan, Miglietta et al. 2009)
Turritopsis sp. 3
Mediterranean (Spain, Miglietta et al. 2009)
Turritopsis sp. 4
W Atlantic (Panama, Miglietta et al. 2009)
Ecology
General:
The hydrozoan Turritopsis dohrnii forms branched hydroid colonies up to 35 mm high. The colonies grow medusa buds which release planktonic medusae. The medusae have separate sexes, and release gametes which fuse to form planulae, which settle to form hydroids. The hydroid stage of this species can be found attached to rocks, docks, pilings and ships’ hulls. A unique feature of the genus is that starved medusae settle to become an undifferentiated cyst, which can grow into a new polyp. This process of transdifferentiation restarts the life cycle, and makes each individual organism potentially immortal (Miglietta et al. 2009).
Food:
Zooplankton
Trophic Status:
Carnivore
CarnHabitats
| General Habitat | Rocky | None |
| General Habitat | Marinas & Docks | None |
| General Habitat | Vessel Hull | None |
| General Habitat | Unstructured Bottom | None |
| Salinity Range | Polyhaline | 18-30 PSU |
| Salinity Range | Euhaline | 30-40 PSU |
| Tidal Range | Subtidal | None |
| Vertical Habitat | Epibenthic | None |
| Vertical Habitat | Planktonic | None |
Tolerances and Life History Parameters
| Maximum Height (mm) | 35 | Bouillon et al. 2004; Schuchert 2004 |
| Broad Temperature Range | None | Warm temperate-Tropical |
| Broad Salinity Range | None | Polyhaline-Euhaline |
General Impacts
No impacts have been reported from introduced populations of Turritopsis dohrnii. However, in Panama Bay, Panama this was one of the seven most abundant hydromedusan taxa (Miglietta et al. 2008).Regional Distribution Map
| Bioregion | Region Name | Year | Invasion Status | Population Status | Vectors |
|---|---|---|---|---|---|
| MED-III | None | 1883 | Crypogenic | Established | |
| MED-II | None | 0 | Crypogenic | Established | |
| MED-VII | None | 0 | Crypogenic | Established | |
| CAR-I | Northern Yucatan, Gulf of Mexico, Florida Straits, to Middle Eastern Florida | 2006 | Non-native | Established | |
| CAR-III | None | 2006 | Non-native | Established | |
| SEP-H | None | 2007 | Non-native | Established | |
| NWP-2 | None | 2003 | Non-native | Established | |
| S190 | Indian River | 2006 | Non-native | Established | |
| MED-V | None | 2003 | Crypogenic | Unknown | |
| NWP-3b | None | 0 | Non-native | Established | |
| PAN_PAC | Panama Pacific Coast | 2007 | Non-native | Established | |
| PAN_CAR | Panama Caribbean Coast | 2006 | Non-native | Established | |
| MED-VIII | None | 0 | Non-native | Established | |
| NEP-VI | Pt. Conception to Southern Baja California | 2010 | Non-native | Established | |
| P040 | Newport Bay | 2010 | Non-native | Established | |
| G260 | Galveston Bay | 2017 | Non-native | Established | |
| SA-II | None | 2020 | Non-native | Established | |
| CAR-VII | Cape Hatteras to Mid-East Florida | 2006 | Non-native | Established |
Occurrence Map
| OCC_ID | Author | Year | Date | Locality | Status | Latitude | Longitude |
|---|
References
Bouillon, Jean; Medel, Maria Dolores; Pagès, Francesc; Gili, Josep-Maria; Boero, Ferdinando ; Gravili, Cinzia (2004) Fauna of the Mediterranean Hydrozoa., Scientia Marina 68(suppl. 2): 5-438Kingon, Kelly; Robertson, D. Ross 2019 The alien Damselfish, Neopomacentrus cyanomos at Trinidad. https://zenodo.org/records/3364568
Marquez, Federico M.; Zabala, Soledad; Bokenhans, Verena; Cumplido, Mariano; Espinosa, Free; Bigatti, Gregorio; Averbuj, Andres (2024) Predation of the invasive green crab Carcinus maenas on the edible snail Buccinastrum deforme, targeted as the most important nearshore marine gastropod fishery from Patagonia, Argentina, Regional Studies in Marine Science 69(103299): Published online
https://doi.org/10.1016/j.rsma.2023.103299
Miglietta, M. P.; Piraino, S.; Kubota, S.; Schuchert, P. (2006) Species in the genus Turritopsis (Cnidaria, Hydrozoa): a molecular evaluation., Journal of Zoological Systematics and Evolutionary Research 45: 11-19
Miglietta, Maria Pia; Rossi, Marco; Collin, Rachel (2008) Hydromedusa blooms and upwelling events in the Bay of Panama, Tropical East Pacific., Journal of Plankton Research 30(7): 783-793
Miglietta, Maria Pia; Lessios, Harilaos A. (2009) A silent invasion., Biological Invasions 11: 825-834
Morri, Carla; Puce, Stefania; Bianchi, Carlo Nike; Bitar, Ghazi; Zibrowius, Helmut; Bavestrello, Giorgio (2009) Hydroids (Cnidaria: Hydrozoa) from the Levant Sea (mainly Lebanon), with emphasis on alien species, Journal of the Marine Biological Association of the United Kingdom 81: 49-62
Schuchert, Peter (2004) Revision of the European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Families Oceanidae and Pachycordylidae, Revue Suisse de Zoologie 111(2): 315-369
Spano, Carlos; Rozbaczylo, Nicolás; Häussermann, Vreni; Bravo, Ricardo (2013) Redescription of the sea anemones Anthopleura hermaphroditica and Bunodactis hermafroditica (Cnidaria: Anthozoa: Actiniaria) from Chile, Revista de Biología Marina y Oceanografía 48(3): 521-534
DOI 10.4067/S0718-19572013000300010